Alternative splicing (AS) is a closely regulated, variable adaptation of the routine RNA modification process of
pre-mRNA splicing. Alternative splicing is a form of
epigenetic mechanism that enables a single
gene to give rise to multiple, differentially spliced versions of a
protein, increasing
complexity without a change in the
genome.
In 1980, a gene called IgM provided the first recognized example of alternative splicing in cells—there were earlier examples in viruses. It has since been demonstrated that cells employ alternative splicing to increase protein diversity toward a variety of biological ends.
An average mammalian gene possesses eight or nine exons—since most human genes undergoing some form of alternative splicing, virtually all of these exons are candidates for elaborate control. The human genome contains 3164.7 million nucleotide bases and only around 25,000 genes, much fewer than the 90,000 proteins that we are estimated to manufacture, and much fewer than prior estimates of 80,000-140,000 expressed sequence tags (ESTs).
Human Genome Project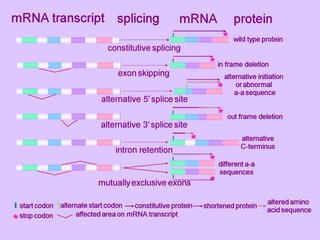
A cell typically
splices a single
mRNA transcript in multiple ways to generate an assortment of
proteins. Alternatively spliced
introns tend to lie between those
exonal segments of a
gene that encode the functional units, or domains, of a protein (
ORF).
Alternative splicing can alter the mRNA product in several ways. (click to enlarge image at left)
At the simplest level, an exon can be removed (
exon skip), lengthened or shortened (alternative
5'AS or
3'AS splicing). Thus, observed mechanisms of alternative splicing alteration include
exon skipping,
intron retention, and the use of an alternative splice
donor or
acceptor site. [
r]
Alternative splicing can affect alternative
promoters, internal exons, or alternative terminal exons, and generates segments of mRNA variability that shift the reading frame, insert or remove amino acids, or introduce a premature termination codon (Fig.
2). The variable segment within the mRNA results from insertion/deletion, or a mutually exclusive swap. The effects on coding potential are an in-frame insertion or deletion, a reading-frame shift, or introduction of a stop codon. mRNAs containing a stop codon >50 nt upstream of the position of the terminal intron are degraded by
nonsense-mediated decay. Therefore, introduction of a premature termination codon into an mRNA by alternative splicing can be a mechanism to down-regulate expression of a gene. Alternative splicing also affects gene expression by removing or inserting
regulatory elements controlling mRNA stability (
NMD or
nonstop decay),
translation, or localization.
Exon skipping is the most frequent
alternative splicing mechanism known in mammals, constituting about 40% of AS events, and as such is a major contributor to mammalian
protein diversity. Exon skipping results in the loss of an
exon in the alternatively spliced
mRNA.
Alternative donor
5'AS or alternative acceptor
3'AS splicing together contribute to 25% of all AS events
conserved between humans and mice (
4).
Intron retention is defined by the presence of a transcript-confirmed intron within a transcript-confirmed exon. Intron retention occurs when introns are not spliced out of the RNA transcript, resulting in the intron(s) being retained within the mRNA as part of an exon. This lengthening mechanism the commonest form of
alternative splicing in plants and lower multicellular organisms.
Regulation of AS:
Alternative splicing depends upon a
splice-site and nearby
enhancer and
repressor sequences—short segments of RNA that couple with
regulatory proteins. It has been estimated that the splicing of a single exon may be promoted by at least three to seven enhancer sequences.
As a result of alternate splicing,
mutations that alter a splice site or a nearby
regulatory sequence can have subtle effects by shifting the ratio of the resulting proteins without entirely eliminating any form.
Regulation of alternative splicing is often temporal or tissue-specific, generating different protein isoforms in different developmental states or different tissues. At the level of the organism, specific isoforms are known to be produced as a consequence of regulation by extracellular
signaling mechanisms. Alternative splicing can allow one gene to generate different proteins in different tissues, providing the diversity encoded within genomes that are smaller than would be expected from gene expression by the organism. Many highly specialized brain proteins arise from differential splicing of genes that are also expressed in other tissues. Cells can even modify splicing in response to changing conditions, and not only can alternative splicing tweak the structure of a single protein, but it may also be a means of regulating entire
pathways.
Recent bioinformatics studies have demonstrated that AS-prone exons can be distinguished from constitutively spliced exons by several features(
1).
1. Divisibility by three of the exon length, which is likely to ensure the preservation of the reading frame in the mRNA (
3).
2. Evolutionary conservation of intronic sequences flanking AS exons (
3–
8). Though not yet fully understood, the unusually high
conservation of the introns that surround AS exons suggests the presence of cis-regulatory elements with regulatory proteins (
5).
Alternative splicing is controlled by the binding of
trans-acting protein factors to
cis-acting sequences within the
pre-mRNA, which result in differential use of splice sites. Since the splice sites do not contain enough information to explain the complex regulation of AS, this fine-tuned mechanism is achieved by multiple weak signals across the exons and introns which are recognized by an extensive number of different proteins (
2). Many such alternative-binding sequences have been identified and are grouped as either enhancer or suppressor elements. These elements are typically short (8-10
nucleotides) and are less
conserved than the
splice sites at exon-intron junctions. The elements are either intronic or exonic, and either enhancers or
silencers depending upon whether they increase or decrease splicing –
ISE,
ISS,
ESE,
ESS.
Members of the
SR (serine rich) splicing factors control alternative splice site recognition by binding to the splicing
enhancer and
inhibitor elements (
ESE,
ESS,
ISE,
ISS). The SR proteins interact with the pre-mRNA substrate and with
snRNP proteins (
small nuclear ribonucleoproteins , pronounced 'snurps'). The role of SR splicing factors in regulating splice site selection is believed to occur in arginine-serine (RS) domain
dependent and RS domain
independent mechanisms.
animation of alternative splicing if this link does not work then click on figure 1 of
alternative splicing link :
life cycle of an mRNA ~ click on Quicktime Q :
HHMI Feature Article on Alternative Splicing :
Artist's conception of ASControlling the Synapse — 49 Proteins at a Time :
The Alternative Splicing Website :
Alternative Splicing DB (ASDB) :
DNA-RNA-ProteinNational Center for Biotechnology Information












































{kind=link}